Презентация на тему "Эволюция иммунитета" подходит как для школьников(8 класс), также и для студентов с биологиеским уклоном. Описание развития иммунитета.
Создайте Ваш сайт учителя Видеоуроки Олимпиады Вебинары для учителей
Презентация на тему"Эволюция иммунитета"
Вы уже знаете о суперспособностях современного учителя?

Тратить минимум сил на подготовку и проведение уроков.

Быстро и объективно проверять знания учащихся.

Сделать изучение нового материала максимально понятным.

Избавить себя от подбора заданий и их проверки после уроков.

Наладить дисциплину на своих уроках.

Получить возможность работать творчески.
Просмотр содержимого документа
«Презентация на тему"Эволюция иммунитета"»
Эволюция иммунитета: введение
Изучение механизмов какого-либо явления у млекопитающих, как правило, сталкивается с определенными трудностями в силу эволюционно сложившейся многофакториальности процессов, лежащих в основе любого биологического феномена.
Обращение к филогенетически менее организованным формам жизни с постепенным переходом от групп, в которых данное явление только зарождалось, к группам, в которых оно усложнилось, помогает изучить отдельные элементы процесса, установить их взаимосвязь и тем самым прийти к пониманию явления. Главные вопросы эволюционной иммунологии связаны с решением проблемы возникновения способности к специфическому антигенному распознаванию (появлению антигенраспознающих рецепторов как молекулярных факторов такого распознавания) и определением путей эволюционного происхождения лимфоцитов - основных участников иммунологических событий.
Существует и другой аспект исследований эволюционного развития иммунной системы. В книге "Целостность организма и иммунитет" М.Бернет (1964) впервые определил иммунитет как ту реакцию организма, которая направлена на поддержание генетической стабильности соматических клеток организма. Подобное определение основано на том представлении, что иммунная реакция развивается не только на экзогенный чужеродный материал, но и на мутационно измененные собственные клетки.
Действительно, спонтанный мутационный процесс в соматических клетках представляет собой неизбежное следствие многоклеточности. Из этого следует, что чем больше соматических клеток у тех или иных филогенетически отличающихся форм жизни, тем больше вероятность генетических нарушений этих клеток и тем эффективнее должен быть контроль за мутационным потоком.
Принимая определение М.Бернета, с неизбежностью следует признать, что иммунитет, взявший на себя функцию контроля за генетической целостностью организма и совершенствуясь на пути своего становления, обеспечил прогресс в мире животных по линии увеличения абсолютного количества соматических клеток эволюционно развивающихся форм жизни. Эту точку зрения поддерживали Н.В.Тимофеев-Ресовский с соавторами (1969), отмечая, что эволюционное изучение какого-либо биологического явления может внести определенный вклад в понимание, и возможно, в изменение теоретических взглядов на течение всего эволюционного развития жизни.
Таким образом, эволюцию специфического иммунитета следует рассматривать не столько как самостоятельное явление исторического развития, сколько как процесс, обеспечивший морфо-функциональный прогресс в мире животных.
Филогенетическое древо взаимных отношений типов животных и иммунитет
Изучение разнообразных видов позвоночных позволяет составить представление об эволюционном развитии иммунной системы вплоть до формирования ее сложноорганизованных механизмов у млекопитающих.
Однако филогенез системы приобретенного иммунитета позвоночных, особенно ее молекулярных основ, остается неясным, несмотря на многочисленые исследования у беспозвоночных.
В то же время изучение беспозвоночных проливает свет на происхождение врожденного иммунитета (например, фагоцитоза ), полностью сформированного у беспозвоночных. Поскольку беспозвоночные чрезвычайно многочисленны и разнообразны (более 95% всех видов животных на Земле), среди них можно найти много удобных объектов для экспериментальных исследований.
Филогенетическое древо - схематизированный образ возможных эволюционных преобразований в мире животных - выступает в качестве матрицы, в ячейки которой закладывается информация о состоянии иммунитета определенной таксономической группы, что позволяет сопоставить формы иммунной реактивности различных групп, в той или иной степени удаленных друг от друга ( рис. 20.1 ).
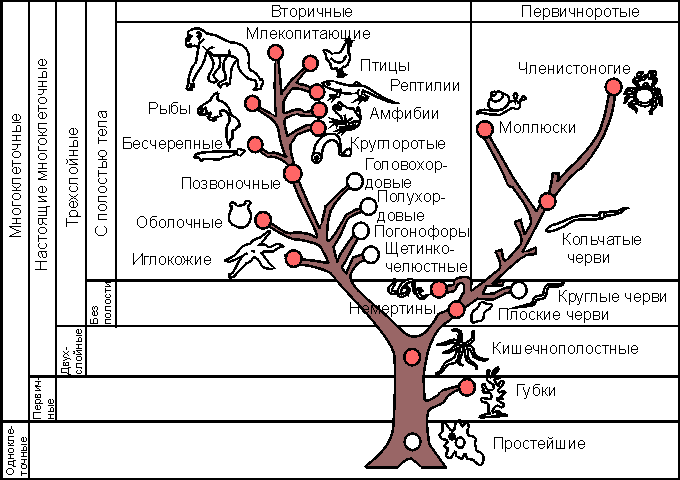
Филогенетическое древо взаимных отношений между основными типами животных.
Красными кружками отмечены типы, у представителей которых изучались те или иные формы иммунитета.
Основные этапы эволюционного развития иммунитета
До середины 60-х годов ХХ-го столетия иммунологи проводили резкую грань между способами иммунной защиты у беспозвоночных и позвоночных животных, считая, что все беспозвоночные, включая хордовых , обладают только неспецифической формой реагирования, тогда как позвоночные животные, начиная с круглоротых , являются носителями и приобретенного иммунитета , свойствами которого являются специфичность в распознавании антигена , индуцибельность и формирование иммунологической памяти .
Подобные различия в иммунной реактивности между двумя группами животных связывали с тимусом позвоночных, отсутствующим у беспозвоночных. Предполагалось, что для беспозвоночных с короткой продолжительностью жизни и значительным потенциалом к самовоспроизведению достаточно неспецифической формы реагирования.
Действительно, неспецифическая клеточная и гуморальная защита у беспозвоночных выражены достаточно хорошо. Однако представления о том, что беспозвоночные лишены каких-либо проявлений специфического иммунного ответа, оказались ошибочными.
В середине 60-х годов была показана способность представителей семейства дождевых червей Eisenia foetida и дождевых червей Lumbricus terrestris к специфическому отторжению аллогеннойткани и формированию кратковременной иммунологической памяти.
Несколько позднее, в 70-е годы, установлено специфическое аллоиммунное отторжение даже у таких низкоорганизованных многоклеточных, как губки и кишечнополостные .
На основании полученных фактов стало ясно, что специфический иммунитет не является прерогативой позвоночных животных. Первые преадаптационные проявления специфического иммунного реагирования возникли, вероятно, очень давно - с момента появления первых многоклеточных около 2 млрд. лет тому назад.
Кроме того, и на это следует обратить особое внимание, выявленные факты позволяют предположить время возникновения предшественников антигенраспознающих рецепторов Т- и В-клеток.
Особое место в изучении вопросов эволюционного развития иммунитета занимает проблема происхождения лимфоцитов - основных участников иммунологических событий.
Вполне вероятно, что лимфоцит как самостоятельный клеточный тип возник исключительно для осуществления клеточной формы иммунного реагирования, и понятия "лимфоидный" и "иммунный" - синонимы для обозначения одной и той же системы организма.
Подобная оценка строится на факте одновременного появления в эволюции нового клеточного типа - лимфоцита - у немертин и кольчатых червей и способности этих животных к специфическому отторжению трансплантата.
Морфологический аспект эволюции лимфоидной системы представляет самостоятельную проблему, в равной мере относящуюся как к гематологии, так и иммунологии.
По представлениям В.Н.Беклемишева (1964), родоначальным клеточным типом, давшим начало внутренней (мезенхимальной) среде организма, был блуждающий амебоцит кишечнополостных . Факт появления амебоцита-макрофага автор оценивал как важное арогенное событие, следствием которого явилось формирование целой системы органов, объединеных в лимфо-миелоидный комплекс. Элементы комплекса в виде различного рода лимфоидных скоплений и узелков уже представлены у беспозвоночных с полостью тела: кольчатых червей , моллюсков , членистоногих , иглокожих , оболочников . Однако своего совершенства лимфо-миелоидный комплекс достигнет у высших позвоночных животных.
Таким образом, обращаясь к лимфоидной системе, следует выделить два основных эволюционных события:
1) возникновение лимфоцита как морфологически обособленного клеточного типа, основнымй свойством которого является иммунологическая функция и
2) вычленение из лимфо-миелоидного комплекса лимфоидной ткани (особое место в которой занимает тимус ) в качестве автономной структуры со своими специфическими (иммунологическими) функциями.
Иммунитет как контролирующий фактор эволюции
Многоклеточность и мутационный риск
Механизмы сдерживания мутаций в организме
Многоклеточность и мутационный риск
По определению, мутационные изменения соматических клеток имеют тот же характер, что и подобные изменения половых клеток. М.Бернет, предполагая, что частота мутаций для половых и соматических клеток имеет один уровень, использует следующий расчет.
Ежедневно у человека вступает в митоз от 1011-1012 клеток. Какое-либо мутационное изменение должно встречаться с частотой 10-6 на репликацию. Из этого следует, что в одной генетической смене клеточного пролиферирующего пула, происходящей ежедневно, должно накапливаться количество мутаций, равное 105.
Если принять, что в одной клетке происходит только одно мутационное событие, то к зрелому возрасту (приблизительно к 27 годам; 10000 дней) в организме человека должно накопиться около 109 мутантных клеток.
Цифра значительно занижена, так как выведена без учета более интенсивных пролиферативных процессов в эмбриональном и раннем постнатальном периодах развития.
Подобный уровень мутационных нарушений слишком велик, чтобы не приносить ущерба сбалансированной системе целого организма. Особенно следует помнить, что некоторые мутации могут привести к интенсивной клеточной пролиферации, значительно превышающей скорость нормальных пролиферативных процессов.
При рассмотрении вопросов о механизмах защиты индивидуального генотипа следует обратить внимание еще на один факт.
Злокачественные новообразования являются основным заболеванием, причину возникновения которого, помимо прочего, связывают с мутационными нарушениями в соматических клетках. И это заболевание имеет статистически достоверное возрастное распределение. Например, наибольшая встречаемость различных форм неоплазм у человека отмечается после 40 лет.
Естественно предположить, что подобное явление не зависит от увеличения частоты мутаций с возрастом. Напротив, так как по мере старения скорость пролиферативных процессов снижается, вероятность ошибок падает. Остается допустить возрастное подавление эффективности механизмов, элиминирующих измененные клетки.
Ясно, что уровень мутационного риска тем выше, чем из большего количества соматических клеток состоит определенный организм.
Тем не менее эволюционное возникновение многоклеточных организмов от одноклеточных объясняют тем, что возможности многоклеточного организма по сравнению с одноклеточным предшественником значительно шире в его борьбе за существование.
Процесс развития многоклеточных шел как по пути увеличения абсолютного количества соматических клеток, так и по пути усиления специализации и дифференцированности отдельных групп клеток. Своего совершенства этот прогрессивный процесс достиг у позвоночных животных.
Однако увеличение числа соматических клеток у эволюционирующих форм жизни шло параллельно с усилением мутационной опасности для клеток тела и, следовательно, всего организма в целом, поскольку мутационный риск является следствием возникновения и развития многоклеточности. Он выражает собой как бы дань за те преимущества, которые получают многоклеточные формы в их борьбе за существование.
На рис. 20.13 показана иллюстрация высказанного положения. Если представить себе гипотетический ряд животных, у которых число возобновляющихся за единицу времени клеток от одного представителя к другому увеличивается на порядок, то в начале ряда будут стоять одноклеточные (предположим, это обыкновенная амеба), а в конце ряда - человек, имеющий до 1014 соматических клеток, из которых ежедневно возобновляется от 1011 до 1012 клеток.
Мутационный риск для животных, организм которых содержит то или иное количество делящихся клеток, будет выражаться произведением частоты мутаций (n) на число возобновляющихся клеток тела (Z):
МР = nZ, где "МР" - величина мутационного риска, а "n" - частота мутаций, равная 10-6.
Следует подчеркнуть, что эта величина выводится только для одной смены клеточной генерации.
Из рисунка видно, что риск мутационного поражения возрастает по мере увеличения количества делящихся клеток.
Так, для животных, имеющих незначительное число возобновляющихся клеток (от 1 до 105), мутационный риск будет величиной отрицательной. Потребуется несколько полных смен клеточных поколений (20-4), чтобы возникла одна мутационно измененная клетка. У животных с большим числом возобновляющихся клеток (от 106 до 1011) мутационный риск представляет собой положительную величину. Уже при смене только части делящихся клеток возникнут мутантные клеточные формы.
Появление многоклеточных стало важным арогенным преобразованием в природе. Быть многоклеточным с обилием хорошо дифференцированных клеток полезно для вида. Разнообразие функций, выполняемых отдельными специализированными клеточными популяциями и взаимная связь между клетками, определяющая организм как единую систему, становится результатом прогрессивной эволюции.
Собственно два показателя: увеличение абсолютного числа клеток и их все увеличивающаяся специализация - являются наиболее демонстративными признаками морфо-функционального прогресса в мире животных. Однако быть эволюционно развитым организмом с астрономическим числом высокодифференцированных клеток также и крайне опасно. Эта опасность кроется в самом факте многоклеточности, в неизбежности сопутствующего мутационного риска.
Ясно, что природа должна была выработать механизм, препятствующий накоплению мутантных клеток.
Рисунок 20.13
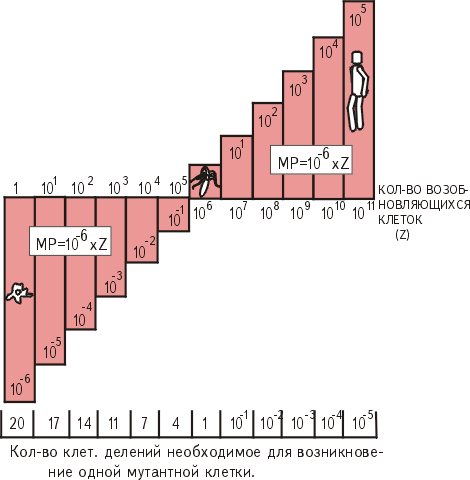
Зависимость величины мутационного риска (МР) от количества возобновляющихся соматических клеток.
Z - число возобновляющихся клеток тела за одну генерацию; 10-6 - частота спонтанных мутаций соматических клеток.
Механизмы сдерживания мутаций в организме
Исходя из представлений, что величина мутационного риска прямо пропорциональна количеству делящихся клеток, следует допустить, что параллельно процессу развития многоклеточности шло формирование механизмов, сдерживающих мутационный поток.
В данном случае необходимо обратить внимание не на внешние биоценотические факторы, а на внутренние, свойственные самому многоклеточному организму.
Без успешного формирования этих механизмов эволюция "застряла" бы на том уровне, при котором количество соматических клеток у какого-либо животного осталось ничтожно мало, а период воспроизведения был короток.
На рис. 20.14 представлено соотношение фактов эволюционного возникновения различных способов иммунного реагирования с уровнем организации в мире животных.
Всего включено пять проявлений иммунитета: фагоцитоз , аллогенная ингибиция , специфическая клеточная форма защиты , реактивность антигенраспознающих рецепторов и продукция иммуноглобулинов .
Способность одноклеточных организмов к фагоцитозу является тем свойством, которое обеспечивает их питание. Фагоцитоз как реакция амебоцитов-макрофагов на чужеродный материал сохранился у всех многоклеточных животных.
Данные по неиммунному распознаванию чужеродности у большинства изученных представителей наиболее просто организованных многоклеточных - губок и кишечнополостных ( табл. 20.3 ), а также демонстрация явления аллогенного ингибирования у млекопитающих позволяют думать, что подобная форма реактивности является общим свойством всех многоклеточных.
Факты зарождения клеточных форм специфического реагирования у простейших многоклеточных и усиление специфического клеточного иммунитета, обусловленного активностью лимфоцитов, у первичноротых и вторичноротых животных определяют рамки, в которых эта форма иммунитета имеет место.
Если верхний предел специфического клеточного реагирования ясен (уровень млекопитающих), то нижний ( губки , кишечнополостные ) выглядит достаточно расплывчатым. Лишь некоторые представители этих таксонов способны к специфическому реагированию с формированием кратковременной иммунологической памяти . Подобные отношения следует отнести к явлению преадаптации элементов клеточной формы реагирования.
На рис. 20.14 квазииммунная форма реагирования отмечена пунктирной линией.
Специфичность клеточной иммунной реакции подразумевает наличие на поверхности эффекторных клеток молекулярных структур, способных к распознавания чужеродного антигенного материала. Несмотря на то, что антигенраспознающие рецепторы у беспозвоночных выявлены только у иглокожих и оболочников , следует предполагать существование этих рецепторов также и у более низкоорганизованных беспозвоночных, способных к специфическому клеточному реагированию. Широкое распространение однодоменных белков суперсемейства иммуноглобулинов ( Thy-1 , бета2-микроглобулина , P0 ) от одноклеточных до высших позвоночных животных вселяет уверенность в том, что какие-то иммуноглобулинподобные антигенраспознающие структуры будут найдены у низкоорганизованных многоклеточных, способных к специфическому реагированию.
И наконец показано, что синтез иммуноглобулинов - компонентов специфического гуморального реагирования - является привилегией позвоночных животных.
Анализ схемы, представленной на рис. 20.13 ясно указывает на необходимость совершенствования иммунных механизмов контроля за процессом эволюционного развития многоклеточных животных.
Конечно, утверждение, что прогресс по линии увеличения количества соматических клеток обеспечивался системой иммунологического контроля за мутационным потоком, было бы более убедительным, если бы удалось провести строгую коррелятивную связь между эволюционно возникающими формами иммунитета и все увеличивающимися размерами носителей этого иммунитета аналогично тому, как это было сделано при рассмотрении иммунитета новорожденных. Однако, отсутствие сведений об истинном количестве воспроизводящихся клеток у представителей разных таксонов и недостаточность знаний о состоянии иммунной реактивности у наиболее просто организованных животных не позволяют сегодня установить абсолютную связь. И тем не менее, можно попытаться провести подобную оценку в гипотетической форме, опираясь на возможные в прошлом пути развития многоклеточности.
На рис. 20.15 в условном графическом виде представлены варианты эволюционных изменений количества возобновляющихся клеток у представителей тех или иных таксономических групп животных. Относительная клеточная величина отображена размером круга (чем больше предполагаемое количество воспроизводящихся клеток, тем больше диаметр круга и наоборот). Условно нарастание многоклеточности обозначено последовательными цифрами 1 - 9. Следует еще раз подчеркнуть, что данная схема полностью умозрительна и указывает лишь на возможные тенденции в эволюционном развитии многоклеточности.
В глубоком геологическом прошлом (очевидно, в архее) эволюция по линии увеличения количества соматических клеток, вероятно, завершилась бы на стадии С4 и не имела бы успеха при дальнейшем увеличении клеточности по причине отсутствия специфического контроля за мутационным потоком. Вместе с тем, при том же уровне многоклеточности форма D4, обладающая определенным видом специфического иммунологического контроля, "обречена" на эволюционный успех. Дальнейшее историческое развитие могло привести как к увеличению абсолютного количества пролиферирующих клеток (линия D-H), так и к различного рода колебаниям по многоклеточности (D-Dn, E-En и т.д.). Это может быть связано с различными условиями существования вида и действием факторов отбора, отличных от иммунных. В результате на уровне современных форм (Dn-Hn) диапазон колебаний многоклеточности велик, а представители с незначительным количеством клеток, но прошедшие предковый путь становления иммунитета, соседствуют с формами, имеющими большее количество соматических клеток, но не обладающими специфическим иммунитетом.
К примеру, наиболее мелкие виды таких высокоорганизованных классов, как птицы и млекопитающие, имеющие полностью сформированные механизмы иммунной защиты, уступают по размерам некоторым видам кишечнополостных, либо не обладающим специфической защитой, либо с очень слабой зощитой.
Иными словами, в тезисе о контролирующей роли исторически развивающихся механизмов специфического иммунитета следует отнести эту роль к начальным этапам становления многоклеточности, к тому исходному периоду, когда определялся сам путь эволюционного развития многоклеточности.
Каково количество соматических клеток, начиная с которого оно не могло бы увеличиваться без параллельного становления специфических форм иммунологической защиты? Исходя из принципов биогенетического закона Геккеля, следует думать, что такое количество незначительно. Уже на самых ранних этапах зародышевого развития позвоночных животных регистрируются признаки Т- и В-систем иммунитета. Возможно, эта величина составляла бы 10 в 6-ой степени воспроизводящихся клеток - тот предел, с которого мутационный риск становится положительной величиной ( рис. 20.13 ).
Таким образом, на эволюцию иммунитета не следует смотреть только как на самостоятельную линию исторического развития, связанную с антиинфекционной защитой; эволюция иммунитета очевидно является процессом, самым тесным образом связаным с эволюцией многоклеточных вообще, и поэтому одним из важных факторов прогрессивной эволюции в мире животных.
Выяснение механизмов, препятствующих мутационному потоку в процессе индивидуального развития, - это в действительности изучение еще одного фактора стабильности в биологии. Без формирования в онтогенезе и эволюционного развития в филогенезе механизмов иммунологического контроля за соматическим мутагенезом жизнь достаточно организованных многоклеточных была бы невозможна.
Рисунок 20.14
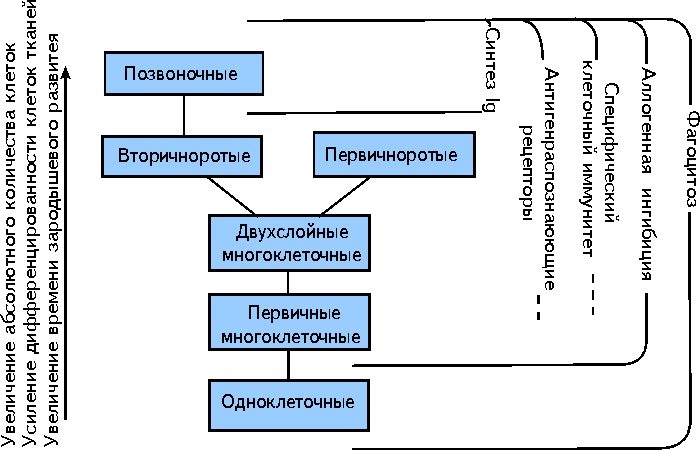
Соотношение между различными формами иммунитета и уровнями организации в мире животных
20.15
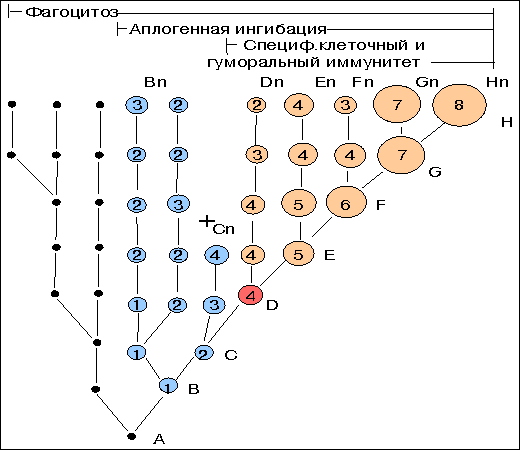
Схема включения иммунных механизмов контроля за эволюцией многоклеточности.
Точки - одноклеточные; кружки 1-9 - условное обозначение количества воспроизводящихся соматических клеток у эволюционирующих многоклеточных животных; B-Bn, C-Cn, D-Dn - направления условного филогенетического развития; Сn - тупиковый путь развития; Dn - многоклеточный организм, с которого начинается филогенетическое развитие специфического иммунитета.
Эволюция иммунитета: заключительный обзор
Оценивая в целом эволюционное развитие иммунной системы, важно учесть следующие выводы.
1. Определяющая форма защиты от инфекционных агентов у беспозвоночных - неспецифическая, обеспеченная в основном активностью амебоцитов-макрофагов и набором гуморальных факторов. Однако даже у низших многоклеточных, таких как губки и кишечнополостные , наблюдается некоторая форма преадаптации к специфическому иммунному реагированию на чужеродный материал, показателем которой является аллотрансплантационное отторжение с формированием кратковременной иммунологической памяти. Блуждающие фагоцитарные амебоциты очевидно становятся обладателями предкового V-гена.
2. К стратегическим успехам развития иммунной системы следует отнести образование у вторичнополостных беспозвоночных первичноротых и вторичноротых ) специализированной клетки - лимфоцита и его поверхностных антигенраспознающих рецепторов .
Появление лимфоцита как основного эффектора иммунного реагирования следует отнести к категории ароморфного преобразования, поскольку это событие определило дальнейшее развитие целой системы организма, без функционирования которой эволюционное формирование многоклеточных было бы невозможно (по Северцеву).
Знаменательна закономерность развития лимфоцитов в линиях первично- и вторичноротых. Между антигенраспознающими клетками этих филогенетически очень далеко отстоящих линий наблюдается не только функциональная связь (распознавание антигена в реакциях трансплантационного иммунитета и цитолиз чужеродной ткани, способность к адоптивному переносу, ответ на Т-клеточные митогены), но и морфологическая, что подтверждает теорию Заварзина о параллельности развития функционально идентичных клеток и тканей.
3. Особое значение в эволюции иммунитета имеет образование тимуса у круглоротых . Именно с появлением этого органа лимфоидная (иммунная) система приобретает самостоятельную роль в жизнедеятельности организма. И это событие следует отнести к еще одному ароморфозу в развитии системы.
4. Возникновение и развитие вариабельности V-генов Т-клеточного рецептора и иммуноглобулинов является еще одной важной вехой на пути становления иммунной системы. Кажется очевидным, что процессы множественных тандемных дупликаций и мутационной дивергенции генов для обоих типов рецепторов шли параллельно.
5. К существенным свойствам эволюции иммунной системы относится и то, что в процессе ее исторического развития появляющийся вновь признак не исключает предыдущий, от которого он произошел. Так например, амебоциты (макрофаги) низших беспозвоночных, дав начало лимфоцитам, сохранились для иммунной системы, взяв на себя функцию презентации антигена в иммуногенной форме и продукцию иммунорегуляторных цитокинов . В целом, специфическая иммунологическая реактивность млекопитающих впитала в себя весь исторический опыт развития иммунных форм защиты - от одноклеточных и низших многоклеточных до высших позвоночных животных.
6. Сопоставление уровней организации в мире животных от одноклеточных до высших многоклеточных, включая позвоночных, с проявлениями различных форм иммунного реагирования позволяет предположить, что иммунитет явился одним из важных факторов прогресса в мире животных по линии увеличения абсолютного количества соматических клеток.
Эволюция T-системы иммунитета: введение
Т-система иммунитета включает тимус , различные субпопуляции Т-лимфоцитов , группу цитокинов , продуцируемых Т-клетками, и эффекторные молекулы - антигенраспознающие Т-клеточные рецепторы (ТКР) . Ясно, что полноценному проявлению этой системы у млекопитающих предшествовал длительный путь эволюционного развития.
Тимус: ключевые моменты деятельности органа
Тимус , как центральный орган иммунной системы, представляет собой эволюционное приобретение позвоночных животных. У всех беспозвоночных он отсутствует даже в зачаточной форме. Его возникновение у примитивных позвоночных было бесспорно ключевым явлением в эволюции иммунитета, и по своей значимости это событие следует отнести к эволюционному процессу, подходящему под определение ароморфоза.
Действительно, образование специальной органной структуры, основное назначение которой - генерализация в онтогенезе Т-клеточного пути развития, значительно повысило эффективность работы всей системы специфической иммунной защиты .
Именно в тимусе формируются основные функционально активные субпопуляций Т-клеток, именно в тимусе медиаторы иммунитета находят свое наиболее эффективное выражение в регуляции созревания Т-клеточного пула, именно в тимусе созданы условия для клоноспецифической экспансии Т-клеток, и, наконец, именно от тимуса зависит заселение периферии эффекторными и регуляторными клетками, принимающими непосредственное участие в иммунном реагировании.
Тимус: эволюция органа у круглоротых
Возникновение зачаточного тимуса в классе круглоротых определило формирование Т-системы иммунитета в качестве самостоятельного морфо-функционального образования, входящего в общую систему специфической иммунной защиты .
У миксин каких-либо морфологических структур, хотя бы отдаленно напоминающих тимус, еще нет. Однако у более совершенного подкласса миног в области жаберных щелей обнаруживается лимфоидная ткань, которую и считают прообразом лимфо-эпителиального тимуса более высокоорганизованных животных. Это единственное достаточно локальное скопление лимфоцитов с относительно высоким уровнем митотической активности, превышающей пролиферативный потенциал данных клеток в периферической крови.
Тимус: эволюция органа у рыб
Хрящевые рыбы в целом как класс характеризуются наличием тимуса, расположенным в головной части вблизи глазных впадин. Показательно, что у таких примитивных хрящевых рыб, как цельноголовые (Holocephali), тимус уже дифференцирован на корковую и медуллярную зоны.
У представителей другого подкласса - пластинчатожаберных (Elasmobranchii; акулы, скаты) - также имеется тимус, причем он, как и у более высокоорганизованных позвоночных животных, подвержен возрастной инволюции.
В классе костных рыб лимфо-эпителиальный тимус представлен не только у настоящих костных рыб (Teleostei), но и у ганоидных, к которым относятся веслонос (Polyodon spatula) и ильная рыба (Amia calva).
В онтогенезе рыб тимус развивается из эпителия одного или нескольких глоточных карманов. У взрослых рыб он расположен под жабрами в области крепления верхнего конца жаберной дуги. Как и у млекопитающих, тимус рыб является поставщиком зрелых клеток для периферических лимфомиелоидных органов и тканей: селезенки, почки, лимфоидной ткани печени. Выселяющиеся из тимуса клетки создают потенциал долгоживущих Т-лимфоцитов периферии, продолжительность жизни которых более 5 месяцев.
Функциональная роль тимуса в иммунных процессах проявляется, в частности, при формировании ответа к аллоантигенам. К примеру, летальное облучение рыб продлевает жизнь трансплантата, а в ряде случаев чешуя вообще не отторгается; в то же время защита области тимуса свинцовым экраном при облучении сохраняет силу иммунного отторжения аллотрансплантата.
В ранние сроки развития иммунного ответа лимфоциты медуллярной зоны тимуса мигрируют на периферию, чтобы принять участие в реакции на антиген. По мере затухания иммунного ответа количество клеток в медуллярной зоне восстанавливается.
Тимус: эволюция органа у амфибий
В онтогенезе тимус у лягушек возникает очень рано - через три дня после оплодотворения в результате инвагинации дорзального эпителия второго глоточного кармана. У личинок на 6-8-й день развития уже сформированы корковая и медуллярная зоны.
В отличие от млекопитающих кора и медулла отделены друг от друга особым клеточным барьером, обогащенным кровеносными сосудами и IgM-продуцирующими клетками. Кора содержит в основном пролиферирующие лимфоциты, окруженные эпителиальными клетками. Кроме того, здесь представлены макрофаги и дендритные клетки. В медуллярной зоне помимо лимфоцитов и клеток стромы изредка встречаются миелоидные клетки, клетки слизистых покровов и пузырчатые дегенерирующие клетки.
Одно из существенных свойств тимуса амфибий состоит в его способности осуществлять положительную селекцию клонов тимоцитов, распознающих чужеродный антиген в комплексе с собственными антигенами главного комплекса гистосовместимости. Эта ситуация гомологична той, которая известна для млекопитающих
Функциональная роль тимуса в иммуногенезе наиболее полно проявляется в опытах с тимэктомией. Удаление тимуса приводит к резкому ослаблению аллотрансплантационной реакции, подавлению ответа в смешанной культуре лимфоцитов (СКЛ), угнетению пролиферации лимфоцитов к Т-клеточным митогенам и снижению гуморального иммунного ответа к Т-зависимым антигенам. При этом реакции, которые развиваются без прямого участия Т-клеток, такие как ответ лимфоцитов к В-клеточным митогенам и антителопродукция к Т-независимым антигенам, не угнетены у бестимусных животных.
Тимус: эволюция органа у рептилий
Тимус рептилий начинает развиваться на самых ранних этапах онтогенеза. Закладка органа происходит в результате инвагинации дорзального эпителия глоточных карманов.
Тимус ящериц, змей, черепах развивается от разных пар глоточных карманов. Например, у ящериц данный орган берет свое начало от второго и третьего, у змей - от четвертого и пятого, у черепах - от третьего и четвертого глоточных карманов. Транзиторные и рудиментарные тимусные почки могут возникнуть также от эпителиальных закладок других глоточных карманов.
У взрослых животных в зависимости от принадлежности к одному из четырех отрядов рептилий наблюдаются анатомические вариации органа как по локализации, так и по числу сформированных долей.
Например, у гаттерии и большинства ящериц имеются две нерасчлененные доли с каждой стороны шеи. Отдельно взятая доля состоит из коры и медуллы и не подразделяется на дольки.
Тимус крокодила представлен вытянутыми четкообразными структурами, которые начинаются у основания черепа, проходят по всей длине шеи и заканчиваются в области сердца. Подобная анатомия тимуса крокодила напоминает морфологию тимуса птиц.
Наиболее структурирован тимус у черепах. Он представлен одной долей с каждой стороны шеи в районе раздвоения сонной артерии. Отдельно взятая доля, в свою очередь, делится на более мелкие дольки с собственной корой и медуллой.
Кора тимуса представлена плотно упакованными малыми лимфоцитами, которые окружены тонкой сетью звездчатообразных эпителиальных клеток. В медуллярной зоне преимущественными клетками являются слабоокрашиваемые эпителиоциты. Количество лимфоцитов в медулле относительно невелико. Дополнительными клетками этой зоны выступают макрофаги, эозинофилы и моноциты.
Как и у всех других позвоночных животных, тимус рептилий подвержен возрастной инволюции, выражающейся в нивелировке различий между корой и медуллой и замещением лимфоидной массы органа соединительной тканью.
Тимэктомия и антилимфоцитарная сыворотка значительно снижают ответ к тимусзависимым антигенам. Это подавление коррелирует с истощением Т-зон в селезенке.
Тимус: эволюция органа у птиц
Тимус птиц представляет собой набор семи пар желез, расположенных на правой и левой сторонах шеи от нижней челюсти до грудной клетки. Нижние доли могут быть погружены в ткань щитовидной железы. В отдельно взятой доле различают кортикальную и медуллярную зоны.
В онтогенезе тимус птиц начинает свое развитие из третьего и четвертого глоточных каналов. Первые лимфоциты в зачатке тимуса появляются на 11-й день эмбриогенеза. Это крупные клетки с диаметром около 11 мкм. В последующие дни большие лимфоциты замещаются клетками меньших размеров. Так, между 11-м и 13-м днями эмбрионального развития преимущественный размер тимоцитов составляет 8 мкм, к 16-му дню основная масса клеток имеет размер около 5,5 мкм. Смена гистологической картины связана с процессами внутритимусной дифференцировки и поэтапного перехода клеток от незрелых предшественников к более зрелым формам.
Как и у млекопитающих, тимус птиц является поставщиком функционально отличающихся субпопуляций Т-клеток .
T-клетки: эволюция и морфология
Лимфоцитоподобные клетки обнаружены у простейших червей и всех целомических беспозвоночных животных. Наиболее полно данного типа клетки изучены у кольчатых червейсем. Lumbricidae.
В очаге отторжения ксенотрансплантата у дождевых червей Lumbricus terrestris и Eisenia foetida скапливаются малые лимфоциты (тип I) и большие (тип II). Именно с этими клетками связан процесс деструкции и отторжения чужеродной ткани. Размер лимфоцитоподобных клеток равен 5-30мкм, и они составляют более 60% от всех клеток целомической жидкости.
Малые лимфоциты представляют собой овальные или сферические клетки с незначительным количеством цитоплазмы. Среди органелл представлены митохондрии, аппарат Гольджи и шероховатый эндоплазматический ретикулум. Кроме того встречаются дополнительные включения в виде прозрачных гранул и темных вакуолей.
Несколько более крупные лимфоцитоподобные целомоциты типа II морфологически подобны клеткам I-го типа. Однако у них более значителен объем цитоплазмы, которая формирует тонкие псевдоподии, и также увеличено количество внутриклеточных включений ( рис. 20.2 ).
В целом лимфоцитоподобные целомоциты L.terrestris обладают структурным сходством с незрелыми лимфоцитами позвоночных животных.
T-клетки: эволюция маркеров и рецепторов клеточной поверхности
Наиболее характерными маркерами и рецепторами Т-клеток млекопитающих являются: Thy-1(маркер всех тимусзависимых лимфоцитов), рецептор к эритроцитам барана, рецепторы к клеточным митогенам ФГА и Кон А, антигенрапознающий T-клеточный рецептор (ТКР) и, конечно, антигены дифференцировки CD4 и CD8 .
Структуры, серологически родственные белку Thy-1 грызунов, представлены на целомоцитах кольчатых червей , моллюсков , оболочников , рыб , амфибий , рептилий , птиц . Более того, их удалось определить даже у бактерий и дрожжей (методом иммуноблотинга).
Из нервной ткани кальмара ( моллюска ) был выделен гомолог Thy-1 млекопитающих, который содержал 84 аминокислотных остатка и включал уникальный для маркера Thy-1 блок из пяти аминокислот.
Логично предположить, что первоначально данный маркер не был связан только с одним типом клеток. Лишь последующие эволюционные преобразования определили его принадлежность Т-клеткам. В данном случае важна не эволюционная динамика, а тот факт, что Thy-1 представлен как на тимусзависимых клетках млекопитающих, так и на целомоцитах первичноротых и вторичноротых беспозвоночных.
Помимо маркера Thy-1 у дождевых червей обнаружен рецептор к эритроцитам барана, свойственный для Т-клеток млекопитающих.
Целомоциты как первичноротых, так и вторичноротых способны отвечать пролиферативным ответом на митогены Т-клеток - ФГА и Кон А.
И наконец, что является крайне существенным, у вторичноротых беспозвоночных (морские звезды, оболочники ) обнаружены гомологи антигенраспознающих рецепторов Т-клеток млекопитающих.
T-клетки: эволюция активности
Очевидно, что о зрелости клеток организма следует судить по тому, насколько эффективно тот или иной класс клеток реализует генетически детерминированные функции и насколько быстро клетки отвечают на сигналы для осуществления запрограммированной активности.
По отношению к Т-клеткам как единой клеточной популяции Т-системы иммунитета показателями полноценности функционального проявления являются:
- эффективность распознавания антигенно чужеродного материала,
- скорость мобилизации (через пролиферацию и дифференцировку) клона или клонов Т-клеток в ответ на внешние антигенные сигналы,
- выраженность регуляторной ( хелперной и супрессорной ) активности,
- способность к сохранению специфического реагирования в виде формирования клеток памятиили сохранению реактивности в системе адоптивного переноса,
- участие в реакциях клеточного взаимодействия,
- продукция молекулярных регуляторов ( цитокинов ) клеточного и гуморального иммунного реагирования.
Ясно, что этот перечень функциональной активности Т-клеток формировался в эволюции постепенно.
Лимфоциты наиболее примитивных позвоночных, каковыми являются круглоротые , хрящевые рыбы и костные рыбы , способны отвечать пролиферативной реакцией на Т-клеточные митогены - ФГА и Кон А, хотя требуют при этом значительно большей дозы, чем лимфоциты более высокоорганизованных позвоночных животных. При этом сам ответ никогда не достигает тех значений, которые известны для Т-лимфоцитов млекопитающих.
В то же время пролифертивный ответ лимфоцитов амфибий и рептилий на Т-клеточные митогены не отличим от ответа Т-клеток птиц и млекопитающих ( табл. 20.4 ).
Реакция в смешанной культуре лимфоцитов (СКЛ) развивается при различиях по антигенам гистосовместимости между взаимодействующими клетками (см." Иммунный ответ и взаимодействие клеток "). Изучение этой реакции в филогенетическом аспекте крайне важно, так как определение в той или иной группе животных способности их лимфоцитов реагировать в СКЛ будет указывать на присутствие в лимфоидной популяции Т-клеток, обладающих поверхностными структурами с антигенраспознающей функцией.
Лимфоциты морских звезд ( иглокожие ) не способны развивать аллогенный ответ в СКЛ. В то же время при различиях между взаимодействующими клетками на видовом уровне регистрируется полноценный ответ.
Отсутствие аллогенного ответа в СКЛ вовсе не означает, что лимфоциты иглокожих ареактивны по отношению к аллоантигенам, поскольку установлено, что животные данного типа отличают "свое" от "чужого" в реакции трансплантационного отторжения. Присутствие у морских звезд поверхностного белка, гомологичного бета-цепи ТКР млекопитающих, также говорит о принципиальной возможности к аллораспознаванию. Ареактивность в алло-СКЛ может быть связана с отсутствием определенного рода костимуляторов, которые существенны в проявлении реакции in virtro и не столь очевидны in vivo.
Круглоротые и хрящевые рыбы развивают лишь слабую реакцию в СКЛ. Сниженная реактивность лимфоцитов данных таксономических групп может быть аргументирована аналогичным образом.
Более высокоорганизованные позвоночные животные - костные рыбы и амфибии развивают вполне выраженную реакцию в СКЛ.
Иммунная система рептилий в морфологическом и функциональном отношениях представлена достаточно полно и сопоставима с системами птиц и млекопитающих . Отражением подобной морфофункциональной зрелости является способность лимфоцитов представителей этого класса развивать полноценный ответ в СКЛ. Однако, если у гомойотермных птиц и млекопитающих подверженность иммунной реактивности сезонным колебаниям выражена слабо или отсутствует (за исключением нескольких видов, впадающих в спячку), то у рептилий напряженность иммунного ответа в значительной степени связана со временем года.
При изучении гетерогенности лимфоцитов у животных рзличных таксономических групп с выделением клеток, выполняющих Т-клеточную функцию, был использован целый ряд экспериментальных моделей. Одна из них связана с изучением типов клеток, реагирующих на комплекс гаптен -носитель. Известно, что у мышей В-клетки отвечают на гаптен, в то время как хелперные T-клетки - на носитель.
Изучение реакции на гаптен-носитель у круглоротых и хрящевых рыб дало отрицательные результаты. При этом лимфоциты костных рыб отличают гаптен от носителя, демонстрируя тем самым гетерогенность по признаку Т-клеток и В-клеток.
У амфибий , как и у костных рыб, воспроизводится феномен гаптен-носитель, в котором именно Т-клетки с хелперной активностью распознают носитель.
Анализ взаимодействия Т- и В-клеток у амфибий обогащен данными о роли антигенов гистосовместимости в клеточной кооперации. Феномен гаптен-носитель был проанализирован, в частности, у лягушки Xenopus laevis в опытах с разделенными на нейлоновой вате Т-клетками, примированными к носителю, и В-клетками, примированными к гаптену. Антитела изотипа IgG, которые являются высокотимусзависимыми, и высокоаффинные IgM-антитела обнаруживались в Т-В клеточной культуре только в условиях идентичности гаплотипов взаимодействующих клеток.
Способностью дифференцировать носитель и гаптен обладают также лимфоциты рептилий , что продемонстрировано в опытах in vitro.
Клеточные взаимодействия при формировании иммунного ответа подразумевают участие клеточных гуморальных факторов, реализующих это взаимодействие. У млекопитающих имеется целый набор Т-клеточных цитокинов , обеспечивающих регуляцию иммунного ответа. Сравнительная иммунология пока не может похвастаться успехами, которые достигнуты при изучении регуляторных белков у млекопитающих, хотя определенные усилия в этом направлении предприняты и касаются они, главным образом, рыб и амфибий. Именно в этих классах позвоночных животных обнаружены цитокины с ИЛ-1 -, ИЛ-2 - и МИФ -подобной активностью. Кроме того, фактор с митогенной активностью выделен из Т-подобных клеток аксиального органа морских звезд.
Эволюция T-системы иммунитета: заключительный обзор
Все беспозвоночные лишены тимуса - места развития Т-клеток. Впервые зачаток тимуса в виде небольшого скопления лимфоидных клеток в районе жаберных щелей появляется у круглоротых .
Возникновение в филогенезе данного морфологического образования бесспорно явилось важным арогенным событием, так как поставило всю систему специфической иммунной защиты на более высокий уровень.
Адаптационное значение данного события легко понять, наблюдая путь, по которому шло эволюционное совершенствование органа от менее структурированной формы низших позвоночных животных к более высокой морфологической и гистологической организации у представителей высокоорганизованных таксонов. Эта тенденция выражается, в частности, в делении органа на доли, дольки, фолликулы Кларка - элементарные гистологические структурные единицы, в наличии телец Гассаля и четком делении паренхимы на корковый и медуллярный слои.
Эволюционное развитие Т-системы связано с возникновением Т-лимфоцита. Знаменательно, что еще до появления специализированного органа, где собственно и происходит "оформление" недифференцированного предшественника в Т-лимфоцит, бестимусные животные ( кольчатые черви , иглокожие , оболочники и др.) уже имеют целомоциты с Т-клеточными свойствами.
О преадаптации целомоцитов беспозвоночных к Т-клеточному пути развития говорит целый ряд фактов.
1. В очагах отторжения алло(ксено)трансплантатов у беспозвоночных представлены клетки, морфологически неотличимые от лимфоцитов позвоночных животных.
2. На целомоцитах представителей многих филумов беспозвоночных обнаружены структуры, родственные наиболее характерному маркеру Т-клеток млекопитающих - антигену Thy-1 .
3. Лимфоциты беспозвоночных, как и клетки позвоночных животных, способны отвечать на Т-клеточные митогены - ФГА, Кон А. Несмотря на то, что этот ответ ниже, чем у высших позвоночных животных, важен сам факт такой возможности. Он говорит о наличии на поверхности лимфоцитов беспозвоночных структур, взаимодействующих с митогенами Т-клеток.
4. Очень существенна информация об экспрессии на поверхности лимфоцитов иглокожих и оболочников отдельных полипептидов или полноценных антигенраспознающих рецепторов, гомологичных соответствующим молекулярным структурам Т-клеток млекопитающих. Эти факты прямо указывают на механизм распознавания чужеродности у беспозвоночных.
5. Лимфоциты иглокожих вступают в реакцию СКЛ на ксеноантигены гистосовместимости, демонстрируя тем самым способность как к распознаванию трансплантационных антигенов, так и формированию ответной реакции - явлениям, свойственным Т-клеткам позвоночных животных.
Этапы гуморального иммунного ответа млекопитающих
У млекопитающих гуморальный иммунный ответ на большинство антигенов ( тимусзависимых антигенов ) включает несколько этапов. Это
- презентация антигена в иммуногенной форме на поверхности фагоцитирующих клеток,
- распознавание антигена Т-лимфоцитами и В-лимфоцитами ,
- антигензависимая дифференцировка В-клеток до продуцирующих антитела плазмоцитов ,
- переключение синтеза одного изотипа антител на другой,
- формирование клеток памяти .
Признаки B-системы в различных таксономических группах
Не все из исследованных групп животных, принадлежащих к различным таксонам, способны к реализации в равной степени антигензависимых процессов, соответствующих этапам гуморального ответа млекопитающих . В табл. 20.5 представлены характерные признаки B-системы иммунитета в различных таксономических группа
Характерные признаки В-системы иммунитета в различных таксономических группах
| Таксон | Интенсивность продукции антител | Память | В-клетки | Плазмоциты |
| Иглокожие | . | - | ± | . |
| Круглоротые: | ||||
| миксины | ± | - | + | - |
| миноги | ± | - | + | + |
| Хрящевые рыбы | + | - | + | + |
| Костные рыбы | + | ± | + | + |
| Амфибии: | ||||
| хвостатые | + | ±* | + | + |
| бесхвостые | ++ | + | + | + |
| Рептилии | ++ | + | + | + |
| Птицы | +++ | + | + | + |
| Млекопитающие | ++++ | + | + | + |
| Таксон
| Основной анти- телопродуцирующий орган | Число изотипов иммуноглобулинов | Выраженность спектротипов | Повышение аффинности антител |
| Иглокожие | аксиальный орган | 1 | . | . |
| Круглоротые: | ||||
| миксины | . | 1 | . | . |
| миноги | жировое тело (взрослые), тифлозоль (личинки) | 1 | . | . |
| Хрящевые рыбы | пронефрос | 1** | ± | - |
| Костные рыбы | пронефрос | 1*** | ± | - |
| Амфибии: | ||||
| хвостатые | . | 3 | ± | - |
| бесхвостые | югулярные тела, костный мозг | 3 | ± | - |
| Рептилии | селезенка, костный мозг | 3 | ± | + |
| Птицы | сумка Фабрициуса | 3 | ± | + |
| Млекопитающие | селезенка | 5 | ++ | ++ |
П р и м е ч а н и я: * - память регистрируется не для всех видов таксона; - двоякодышащие рыбы имеют два или три изотипа; -аксолотль имеет два изотипа иммуноглобулинов.
Эволюция B-системы у иглокожих
Возможно, что первые признаки В-клеток появились у иглокожих . Клетки аксиального органа морской звезды Asterias rubens, прилипающие к нейлоновой вате при фракционировании, способны отвечать на В-клеточные митогены (ЛПС, экстракт из Nocardia opaca). Культура этих клеток в условиях повторной стимуляции гаптеном ТНФ продуцирует антителоподобный фактор, специфичный к соответствующему гаптену.
Насколько эти клетки отвечают требованиям В-лимфоцитов, пока неясно
Эволюция B-системы у круглоротых
Лимфоидная ткань первичных позвоночных животных - круглоротых - представлена слабо.
Например миксины не имеют дискретных морфологически хорошо обособленных и самостоятельных истинно лимфоидных образований. Мононуклеарные клетки лимфоидного типа встречаются в мышечной ткани глоточной и заднежаберной областей, в фолликулах кишечника, паренхиме пронефроса, крови.
С помощью моноклональных антител в периферической крови тихоокеанской миксины выявлено около 65% лимфоцитов с поверхностным иммуноглобулином (sIg) - явным характерологическим признаком В-клеток. Однако у этих животных отсутствует способность формировать плазмоциты, активно продуцирующих антитела - завершающую клеточную форму в гистогенезе В-клеток.
В то же время более высокоорганизованные представители этого класса - миноги - демонстрируют некоторый успех в развитии лимфоидной ткани, в целом, и В-системы, в частности.
Основным органом лимфомиелопоэза взрослых миног является жировое тело - морфологически оформленное структурное образование, расположенное над хордой ( рис. 20.11 ).
У личинок функцию лимфомиелопоэза выполняют тифлозоль (спиральный клапан) - прообраз селезенки, и почка, которые в процессе метаморфоза резорбируются, а их функция переходит к жировому телу (зачатку костного мозга протопозвоночной дуги). В тифлозоле личинок и жировом теле взрослых особей обнаружены плазматические клетки - продуценты иммуноглобулинов (табл. 20.5 ).
Эволюция B-системы у рыб
У хрящевых рыб (акулы, скаты) лимфоциты с поверхностным иммуноглобулином и плазматические клетки представлены в пронефросе, селезенке, лимфо-миелоидном органе Лейдига, связанным с пищеводом, эпигональном органе, расположенным вблизи гонад. В эмбриогенезе первым иммуноглобулин-продуцирующим органом является печень.
У костных рыб около половины лимфоцитов периферической крови обладают поверхностным иммуноглобулином, т.е. являются В-клетками. У этого класса позвоночных животных нет функционально активного костного мозга, сумки Фабрициуса, лимфатических узлов. Функцию первичного органа для В-клеточного пути развития и источника стволовых элементов для лимфоидных и миелоидных клеток выполняет пронефрос ( табл. 20.5 ).
Эволюция B-системы у амфибий
Амфибии представляют собой класс, который делает определенный прорыв в совершенствовании специфического иммунитета.
Связано это, безусловно, с их переходом от водного образа жизни к наземному и, соответственно, с необходимостью иметь дополнительные возможности зашиты от новой группы инфекционных агентов воздушной и почвенной среды. Прогресс в развитии иммунной системы касается, в первую очередь, бесхвостых амфибий, которые в эволюционном развитии становятся обладателями функционирующего костного мозга и лимфатических узлов.
Процентное распределение В-клеток в различных органах лягушек выглядит следующим образом: югулярные тела - 50%, костный мозг - 14%, кровь - 14%, селезенка - 10%, тимус - 1%.
Основным источником В-клеток у лягушек, как и у млекопитающих, является костный мозг. Именно переход к наземному образу жизни, потребовавшему развития конечностей, определил костномозговую ткань как существенное или доминирующее место в преимущественном развитии В-клеточного ростка дифференцировки ( табл. 20.5 ).
Эволюция B-системы у рептилий
Лимфоидные органы и ткани у рептилий достаточно развиты. Помимо специализированных и морфологически хорошо обособленных органов - тимуса, селезенки, костного мозга - лимфоидные скопления встречаются в слизистой кишечника, клоаке, осевом регионе.
Наиболее активным органом в продукции антител, как и у млекопитающих, является селезенка. В селезенке представлено около 50% В-клеток, в периферической крови - 25%, в костном мозге - 20% и в тимусе - менее 1%.
У зародышей большое количество В-лимфоцитов выявлено в печени, которая, очевидно, является главным органом, где происходит дифференцировка В-клеток в раннем онтогенезе (табл. 20.5 ).
Эволюция B-системы у птиц
Сумка Фабрициуса в клоаке, селезенка, железы слепой кишки и железа Гардеотана - органы, где у птиц реализуется антителопродукция. Среди них сумка Фабрициуса занимает доминирующее место. Однако представление о том, что данный орган является первичным в формировании В-клеток, пересматривается ( табл. 20.5 ).
Эволюция изотипов иммуноглобулинов у иглокожих
По устоявшемуся мнению, способность к синтезу антител является привилегией позвоночных животных.
Тем более интересны данные, полученные французским исследователем Леклером ( Leclerc) при работе с иглокожими - морскими звездами. Ему и его соавторам удалось выявить антигенспецифический индуцибельный фактор, который продуцируется клетками аксиального органа. Антителоподобный продукт представляет собой тетрамер с мол. масой субъединицы в 30 кД и всей молекулы - в 120 кД.
Если эти исследования подтвердятся, они будут иметь особый интерес, поскольку иглокожие находятся в одном подразделе с позвоночными животными и эволюционно предшествовали им.
Эволюция изотипов иммуноглобулинов у круглоротых
В классе круглоротых , происшедших от бесчелюстных щитковых, первые находки которых относятся к верхнему ордовику (около 450 млн. лет назад), впервые регистрируются иммуноглобулины.
Иммуноглобулины обнаружены как в подклассе миксин, так и в подклассе миног.
У миксин иммуноглобулин имеет высокомолекулярную форму - около 1 млн. дальтон - и представлен комплексом нековалентно связанных субъединиц с мол. массой каждой от 17 до 33 кД. При этом доминирует компонент с мол. массой в 22 кД, и по этому показателю он подобен легким цепям иммуноглобулинов высших позвоночных животных.
Представители более высокоорганизованного подкласса круглоротых - миноги - обладают иммуноглобулином с мол. массой около 180 кД. Помимо мономерной формы в незначительном количестве представлена более комплексная форма этого иммуноглобулина ( рис. 20.12 ).
Эволюция изотипов иммуноглобулинов у рыб
У хрящевых рыб (акул) обнаружены иммуноглобулины двух молекулярных форм, относящихся к одному и тому же изотипу - с высокой (900 кД) и низкой (189 кД) мол. массой. Как и у млекопитающих, пентамерная форма иммуноглобулина акул объединена в единую молекулу J-цепью .
Помимо чисто "внешнего" сходства между иммуноглобулинами двух далеко отстоящих классов позвоночных животных имеется гомология и по аминокитслотной последовательности.
В классе костных рыб наблюдается определенная гетерогенность молекулярных форм иммуноглобулинов. Однако доминирующими по видам формами являются тетрамер и мономер, относящиеся к одному и тому же изотипу IgM.
В этом же классе впервые в филогенезе возникает дифференцировка иммуноглобулинов по изотипам у двоякодышащих рыб ( рис. 20.12 ).
Эволюция изотипов иммуноглобулинов у амфибий, рептилий, птиц
Для амфибий , рептилий и птиц характерна гетерогенность по иммуноглобулинам.
При этом эволюционная преемственность с млекопитающими установлена только для IgM . Филогенетическая связь других изотипов иммуноглобулинов млекопитающих с изотипами нижестоящих филумов не установлена ( рис. 20.12 ).
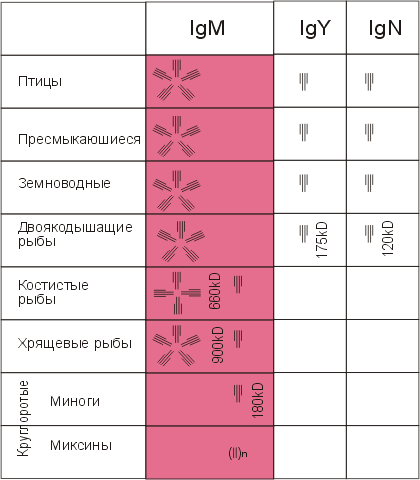 Рис. 20.12. imm_gal
Рис. 20.12. imm_gal Изотипы иммуноглобулинов у позвоночных животных.
Эволюция иммунитета: заключительный обзор
Оценивая в целом эволюционное развитие иммунной системы, важно учесть следующие выводы.
1. Определяющая форма защиты от инфекционных агентов у беспозвоночных - неспецифическая, обеспеченная в основном активностью амебоцитов-макрофагов и набором гуморальных факторов. Однако даже у низших многоклеточных, таких как губки и кишечнополостные , наблюдается некоторая форма преадаптации к специфическому иммунному реагированию на чужеродный материал, показателем которой является аллотрансплантационное отторжение с формированием кратковременной иммунологической памяти. Блуждающие фагоцитарные амебоциты очевидно становятся обладателями предкового V-гена.
2. К стратегическим успехам развития иммунной системы следует отнести образование у вторичнополостных беспозвоночных первичноротых и вторичноротых ) специализированной клетки - лимфоцита и его поверхностных антигенраспознающих рецепторов .
Появление лимфоцита как основного эффектора иммунного реагирования следует отнести к категории ароморфного преобразования, поскольку это событие определило дальнейшее развитие целой системы организма, без функционирования которой эволюционное формирование многоклеточных было бы невозможно (по Северцеву).
Знаменательна закономерность развития лимфоцитов в линиях первично- и вторичноротых. Между антигенраспознающими клетками этих филогенетически очень далеко отстоящих линий наблюдается не только функциональная связь (распознавание антигена в реакциях трансплантационного иммунитета и цитолиз чужеродной ткани, способность к адоптивному переносу, ответ на Т-клеточные митогены), но и морфологическая, что подтверждает теорию Заварзина о параллельности развития функционально идентичных клеток и тканей.
3. Особое значение в эволюции иммунитета имеет образование тимуса у круглоротых . Именно с появлением этого органа лимфоидная (иммунная) система приобретает самостоятельную роль в жизнедеятельности организма. И это событие следует отнести к еще одному ароморфозу в развитии системы.
4. Возникновение и развитие вариабельности V-генов Т-клеточного рецептора и иммуноглобулинов является еще одной важной вехой на пути становления иммунной системы. Кажется очевидным, что процессы множественных тандемных дупликаций и мутационной дивергенции генов для обоих типов рецепторов шли параллельно.
5. К существенным свойствам эволюции иммунной системы относится и то, что в процессе ее исторического развития появляющийся вновь признак не исключает предыдущий, от которого он произошел. Так например, амебоциты (макрофаги) низших беспозвоночных, дав начало лимфоцитам, сохранились для иммунной системы, взяв на себя функцию презентации антигена в иммуногенной форме и продукцию иммунорегуляторных цитокинов . В целом, специфическая иммунологическая реактивность млекопитающих впитала в себя весь исторический опыт развития иммунных форм защиты - от одноклеточных и низших многоклеточных до высших позвоночных животных.
6. Сопоставление уровней организации в мире животных от одноклеточных до высших многоклеточных, включая позвоночных, с проявлениями различных форм иммунного реагирования позволяет предположить, что иммунитет явился одним из важных факторов прогресса в мире животных по линии увеличения абсолютного количества соматических клеток.








Полезное для учителя




Распродажа видеоуроков!

1900 руб.
2930 руб.

1900 руб.
2930 руб.

1860 руб.
2860 руб.

1860 руб.
2860 руб.
ПОЛУЧИТЕ СВИДЕТЕЛЬСТВО МГНОВЕННО

* Свидетельство о публикации выдается БЕСПЛАТНО, СРАЗУ же после добавления Вами Вашей работы на сайт
Удобный поиск материалов для учителей
Проверка свидетельства